『オックスフォードにおける構造生物学』
University of Oxford
Wellcome Trust Centre For Human Genetics
池水 信二
オックスフォード大学の構造生物学
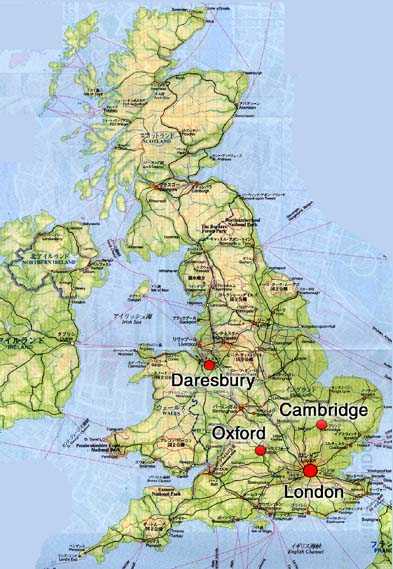
オックスフォードは、ロンドンから北西約100Kmに位置する英国で一番古い学園都市である。オックスフォード大学で一番古いユニバーシィティ-カレッジは、昨年750周年を迎えた。オックスフォード大学のタンパク質結晶学の歴史は古く、Dorothy Crowfoot Hodgkin教授がインスリンの構造解析をしていた。残念ながらインスリンの構造解析には時間がかかり、蛋白質構造解析の第一号にはなれなかった。しかし、彼女はビタミンB12の研究が認められて、1964年にノーベル賞を受賞されている。
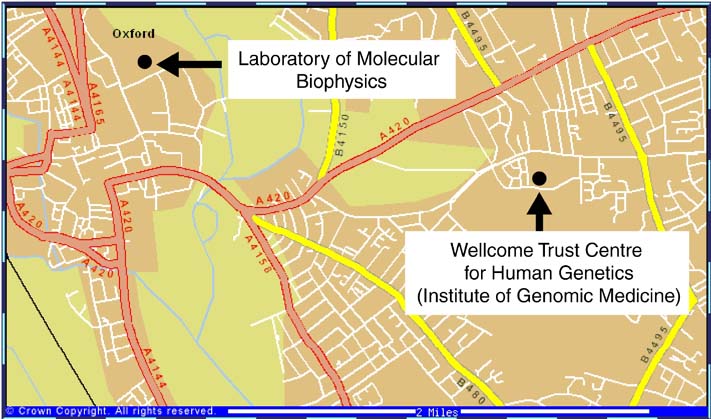
オックスフォード大学には、我々のWellcome Trust Centre for Human Genetics (WTC)にDavid Stuart教授, Yvonne Jones教授, David Stammer博士の3グループ,Laboratory of Molecular Biophysics (LMB)にLouise Johnson教授のグループ(新たに幾つかのグループが形成されつつある)、Dunn School of PathologyにDale Wigley博士等の蛋白質結晶学グループがある。Johnson教授はキナーゼや細胞周期をコントロールする蛋白質、Wigley博士はDNA結合蛋白質の研究を行っている。Oxford Centre for Molecular Science (OCMS)のChristopher Dobson教授は様々な手法を用いて蛋白質のホールディングの研究を、Dept. of BiochemistryのIan Campbell教授はNMRを用いて細胞接着分子やシグナル伝達に関わる蛋白質の研究を行っている。2000年1月にEMBL-Heidelbergから移ってきたWTCのStephen Fuller博士は、電子顕微鏡(EM)を用いてウィルスの構造解析を行っている。
WTC構造生物学グループ
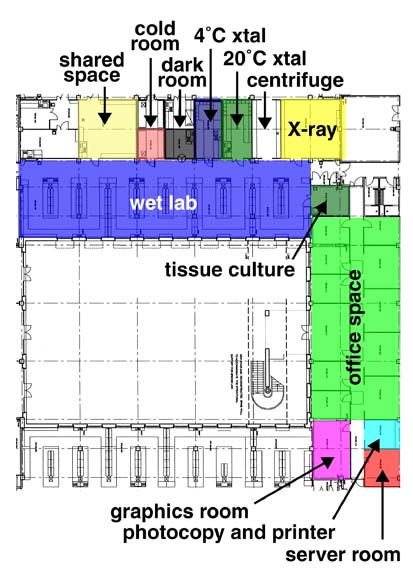
私は1996年5月に日本学術振興会特別研究員としてStuart教授とJones教授のグループに加わった。当時彼らは LMBで研究していたが、昨年6月WTCの完成と同時に、Stuart教授はWTCの構造生物学部門のdirectorとして異動した。WTCは、財団Wellcome Trustの出資によりできた研究所で、OxfordのHeadingtonにあるChurchill Hospitalに面して建てられている。彼の異動に伴いJones教授とStammers博士のグループもLMBからWTCへ異動した。WTCには大きく分けて構造生物学部門とHuman Geneticsの2部門がある。Human Geneticsには現在25のグループがあり、更にノックアウトマウスを用いた研究を行うグループ等が新たに加わる予定である。構造生物学部門は、Stuart教授, Jones教授, Stammers博士の3つの結晶解析グループとFuller博士のEMグループがある。WTC構造生物学部門のコンピュータ管理者としては、bobscriptの作者Robert Esnouf博士がいる。さらにポスドク, PhDコースの学生やテクニシャンを入れると総勢約40名になる。
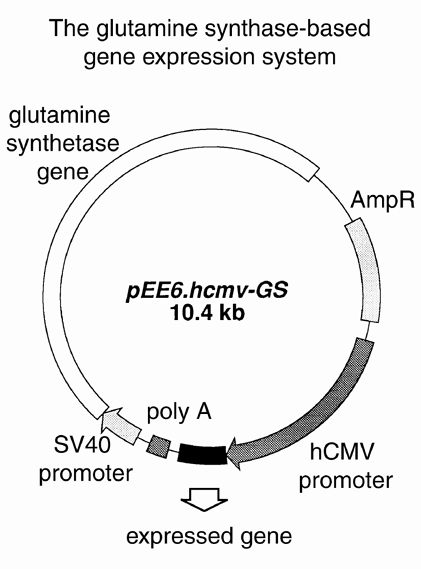
我々は蛋白質を大腸菌,昆虫細胞,CHO細胞を用いて発現させている。昆虫細胞,CHO細胞を用いた場合、post-docや学生が発現系を作成し、細胞培養はテクニシャンによってなされる。蛋白質の精製はpost-doc等が行っているが、精製をするテクニシャンが構造生物学部門に加わったので、今後はテクニシャンによって行われるようになる。
構造生物学部門のコンピュータは、Origin200(dual R10000 225MHz) 2台をホストにユーザー領域, 生データ保管領域として(20*18GB)+10*36の計720GBのRAIDシステムがある。他にサーバーとしはCompaq DS10 (dual ev6 500MHz)が2台ある。グラフィックコンピュータとしてSGI Indigo2が5台, O2が10台, Octane(dual R12000)が1台(同等のものが更に1台増設される)、VW320が7台やDEC PW433auが1台、PCが19台(2台はMar Image Plateのコントローラとして)ある。バックアップ用装置としてStorageTek Timber Wolf[40*DLT7000(70GB)]が導入されており、自動的にバックアップをしている。
実験室系測定装置としては、X線発生器にRIGAKU RU300、光学系にOsmic Multilayer opticsを用いて集光し、Mar Image Plate(300mm又は345mm)を使用して読み取りを行っている。 測定されたイメージは、RAID上に記録され、逐次DLTテープ上に保管される。
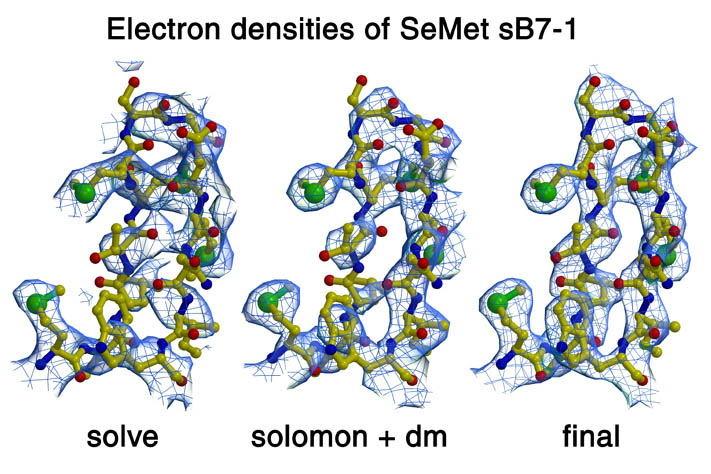
放射光での測定は、Daresbury, ESRFやAPSで行っている。Daresburyは、2.0 GeVのリングで、タンパク質用stationとして7.2、9.5、 9.6,14.1と14.2がある。この中で9.6,14.1と14.2は、ウィグラーからの光を使用しており、ADSCのCCD Quantam-4又はQuantum-4Rとの組み合わせにより比較的早く測定することが出来る。ESRFは6GeVの大変安定したリングで、タンパク質結晶用のstationは、ID9 (Laue,30%を蛋白結晶用に),ID13 (Micro-focus,30%を蛋白用に),ID14 [EH1(単色),EH2 (単色),EH3 (単位格子の大きなものも測定可能) & EH4 (MAD,高輝度), BM14 (MAD) とID29 (MAD,2000年から?)がある。これらのstationはMar CCD 133mm, Mar CCD 165mm又はADSC Quantum-4が設置されている。APSは、独自に開発された3*3 CCDと挿入光源の組み合わせで、振動角1度の振動写真で2-3秒の露光+CCDの読み取り・保存で3秒(ADSC Quantum-4では、読み取り・補正・保存に約10秒要する)の計5-6秒で測定できる。測定された回折イメージは、全てプログラムパッケージ・HKL2000 (現在のバージョンは、1.96.3)により処理している。構造解析用パッケージとしては、CCP4,CNSを主に使用している。分子置換では、CCP4バージョンのAMoRe,MolRep (CCP4 Ver.4.0からパッケージに加えられた),GLRF,CNS,X-PLOR等を使用している。AMoReは、計算時間が比較的短く計算出来るようにアルゴリズムが書かれており、非対称単位に1分子だけの場合はexamplesに含まれている自動化シェルスクリプトを使用して1度のジョブで解を得ることが出来る。MolRepはより自動化されたプログラムで、非対称単位中に複数分子存在する場合に適している。入力で解析する分子数を入力するだけで、一度のジョブで複数分子探すので楽で良い。これらのプログラムをCCP4 GUIを用いて使用した場合、結果もGUIを用いてグラフィカルに表示可能である。これらで解析出来ない場合は、X-PLORやCNSを用いて行う。更にGLRFで全ての回転関数の領域についてトランスレイションサーチを行うことにより解析出来る場合があるが、計算時間を消費するのであまりお勧めではない。同型置換法や多波長異常分散(MAD)法を用いる場合、解析する重金属の数が少ない場合はパターソンを解釈して位置を求めるが、数が多い場合はSHELX,SOLVE,SnB (Shaken-and-Bake)を用いて行っている。我々グループでは、最高50のSe原子の位置をSnBを用いて決定しMAD法により構造解析した例がある。この場合、アノーマラスパターソン又は2波長間(f'の差が大きい)差パターソンでは、SnBを用いても約20のSe原子位置しか求めることが出来なかった。プログラムGAP (General Average Program;Jonathan Grimes博士とDave Stuart教授により書かれた)を用いて2パターソンをスケール及び平均化して、このパターソンを用いてSnBを使用して50原子全ての座標を求めることが出来た。さらにSHARPを用いてSe原子位置の精密化及び初期位相の計算、SOLOMONを用いた電子密度図の改良により解析を行った。蛋白質の構造の精密化は、主にCNSを用いている。現在グラフィックプログラムとしては、Oを使用している。CCP4でもグラフィックプログラムを作成する計画があり、我々のコンピュータ管理者のEsnouf博士は開発計画メンバーである。
研究紹介
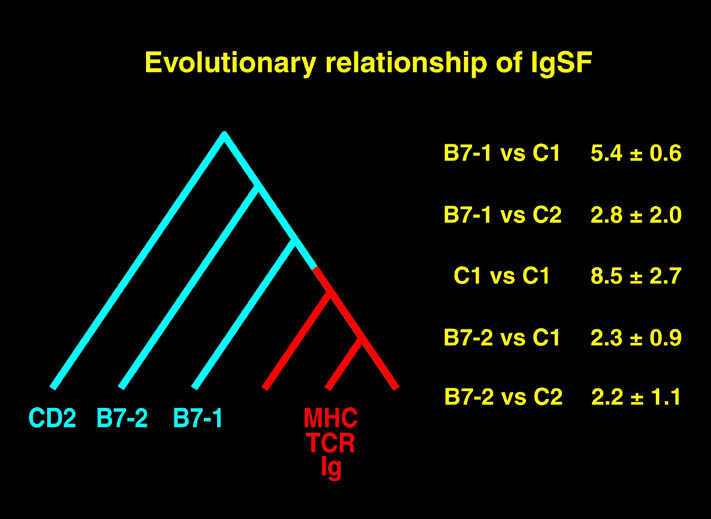
Stuart教授はウィルス,ウィルス蛋白質の構造解析を中心に研究を行っている。更に免疫系のMHCや細胞接着分子の研究をJones教授と共同で行っている。Jones教授は、その他にサイトカインやサイトカイン受容体の研究も行っている。Stuart教授とJones教授の共同で行われた最近の研究には、MHC-CD8複合体,Gelsolin,HIV-1逆転写酵素-阻害剤複合体,HLA-E, Killer cell immunoglobulin-like receptor (KIR),TRAIL-DR5複合体,Fibronectinのヘパリンとインテグリン結合ドメイン,LFA-3やB7-1等がある。Stuart教授グループ単独で行われた研究としては、BTV, BTV10, FMDV-heparan sulfate complex, Histamine結合タンパク等がある。Jones教授のグループでは他にGP130, Sialoadhesion molecule-阻害剤複合体,MHC-糖修飾を受けたペプチド複合体等の研究がある。
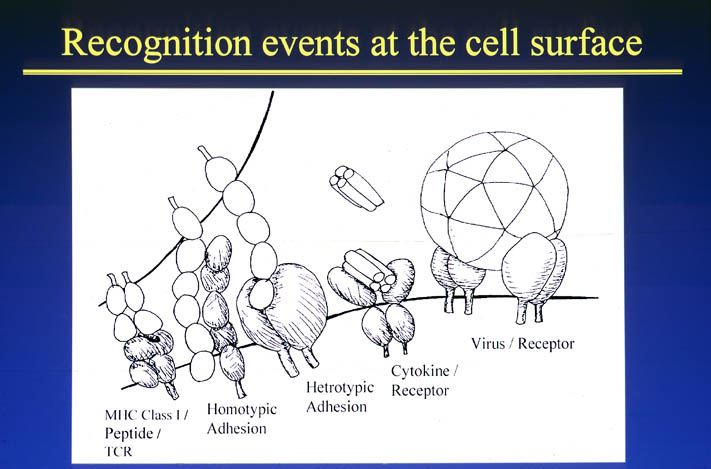
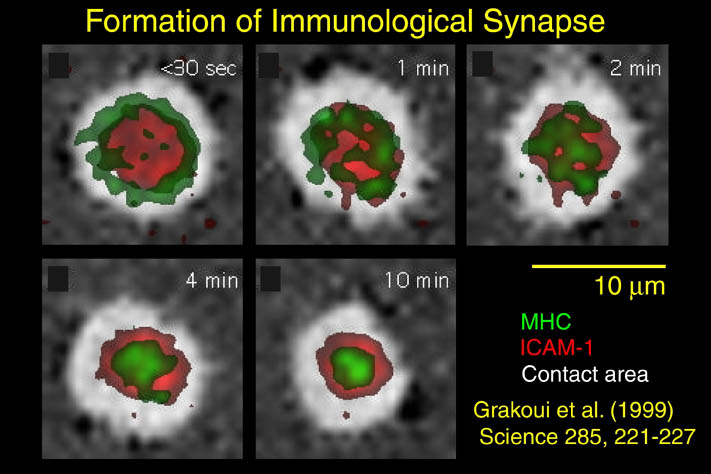
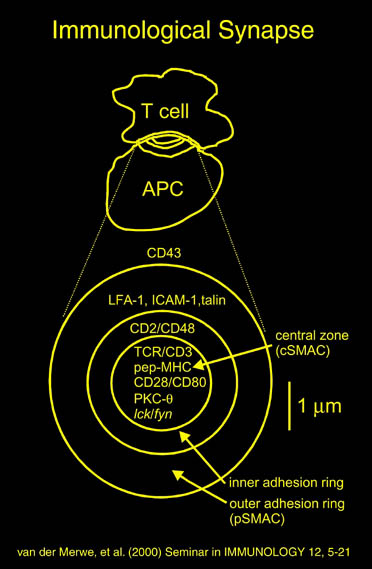
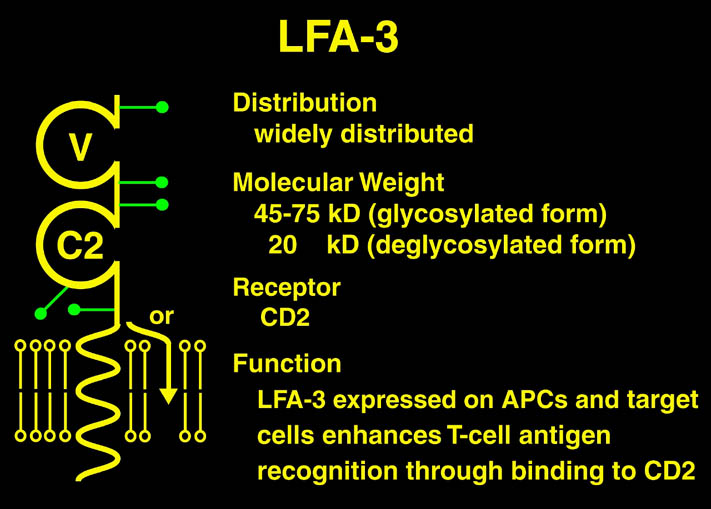
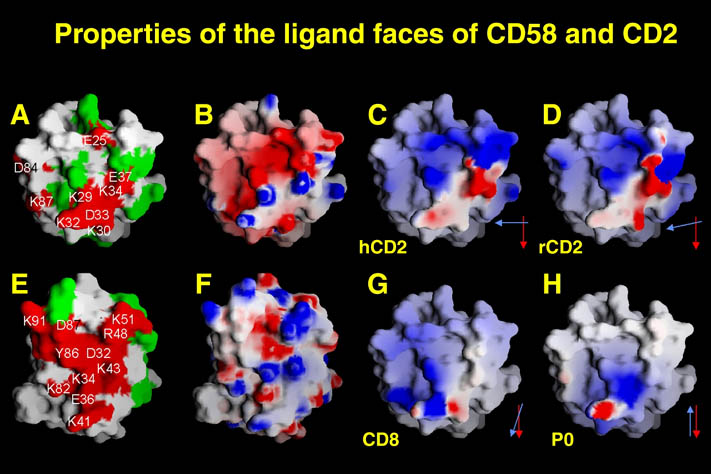
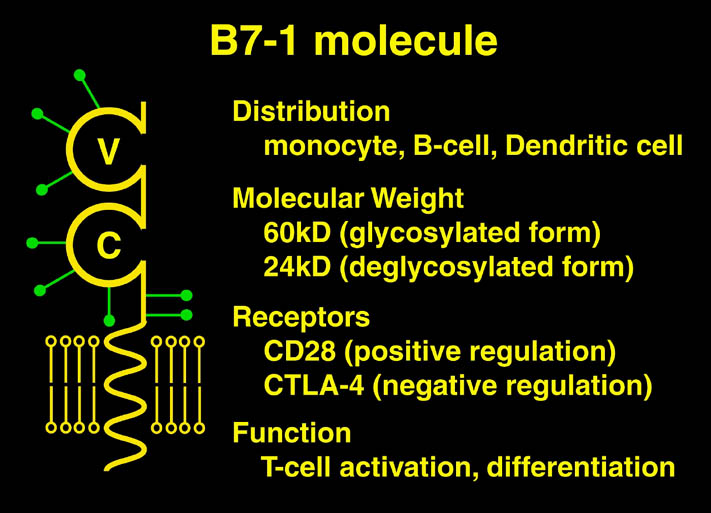
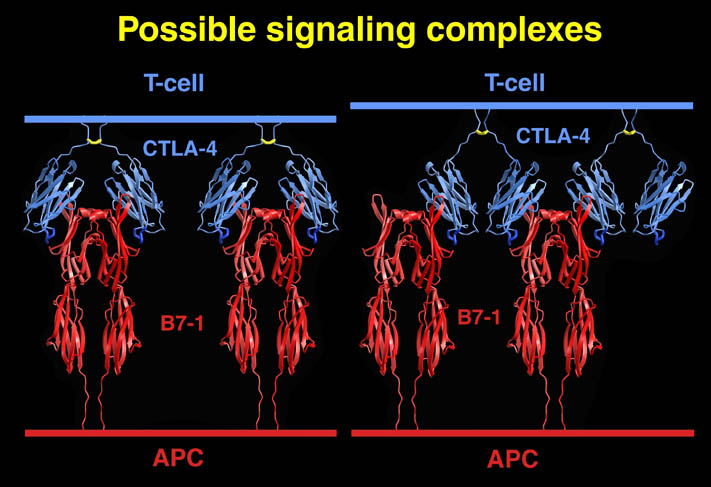
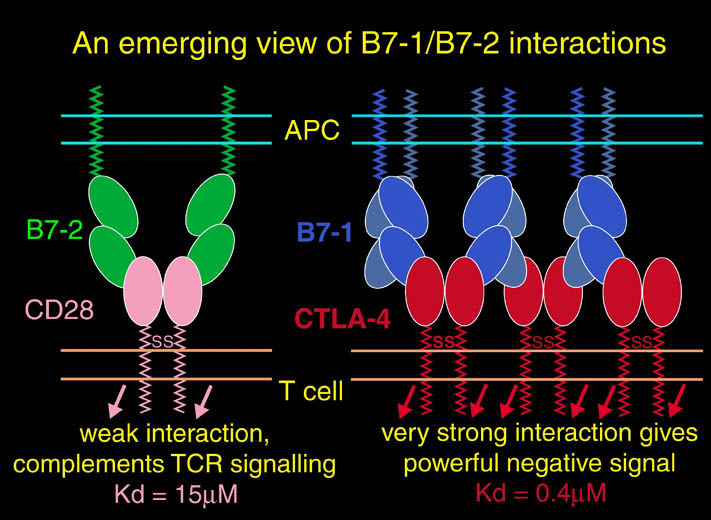
私が解析を行ったものの中からB7-1とLFA-3という免疫系の蛋白質を紹介する。免疫反応にはT細胞の活性化が必要であり、T細胞の活性化には2つのシグナルが必要である。シグナル1は抗原提示細胞(APC)上の主要抗原組織適合複合体(MHC)とT細胞上のT細胞受容体との結合の結果起こる抗原特異的なものである。この結合はT細胞上のCD2とAPC上のLFA-3の結合により増強される。シグナル2は、APC上の補助刺激分子B7-1又はB7-2とT細胞上のCD28の結合の結果起こる抗原非特異的なものである。このシグナル2はリガンドB7分子と受容体CTLA-4の結合により阻害される。シグナル1のみの存在下ではMHCに提示さている抗原ペプチドは自己抗原として認識され、そのT細胞は無反応状態になる。シグナル2のみの場合は、T細胞に変化が起こらない。又、CTLA-4 Fcを用いてB7分子とCD28の結合を阻害することにより、移植片拒絶反応を抑制することも知られている。
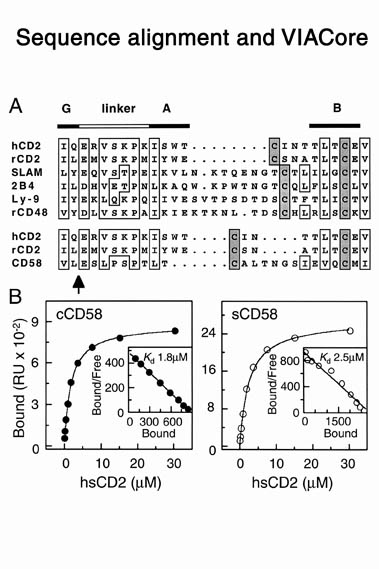
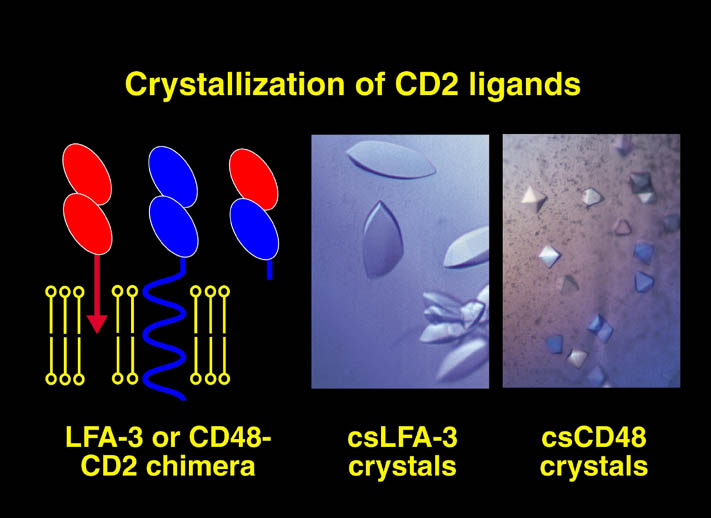
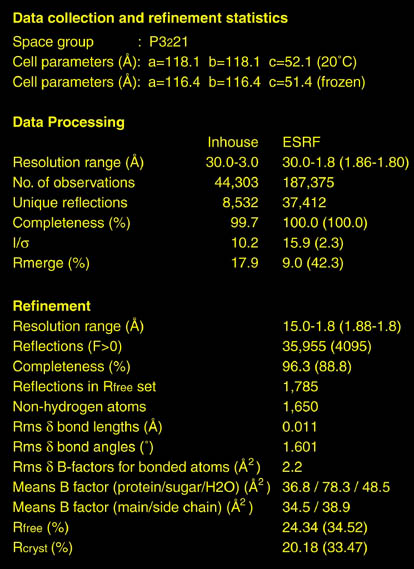
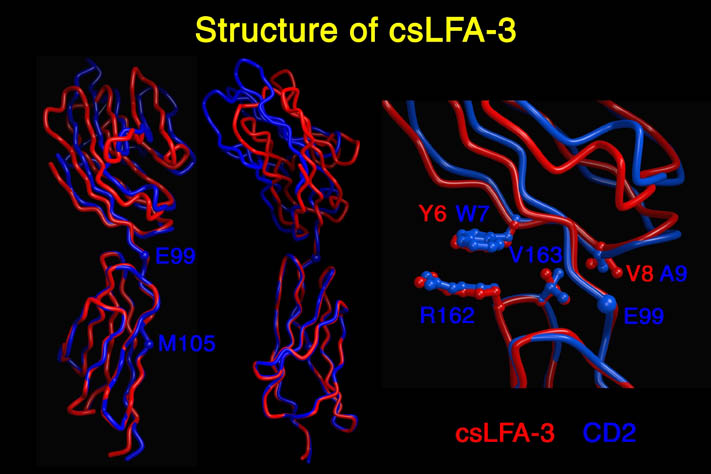
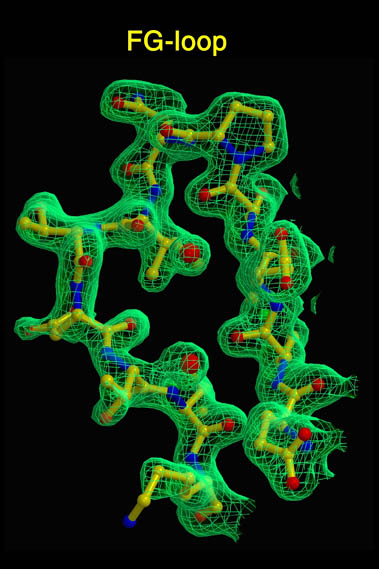
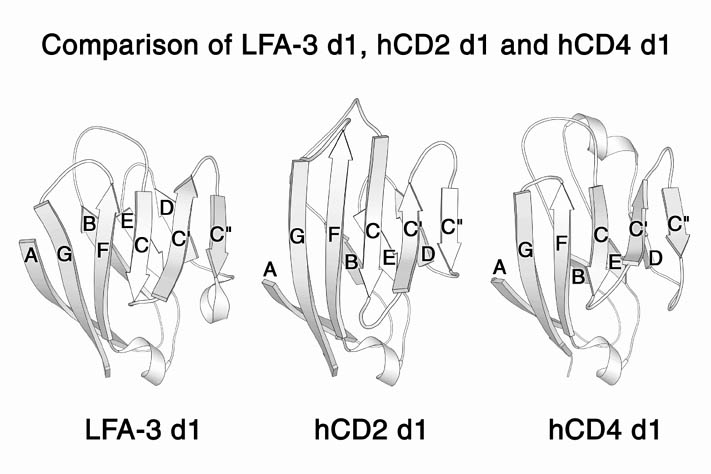
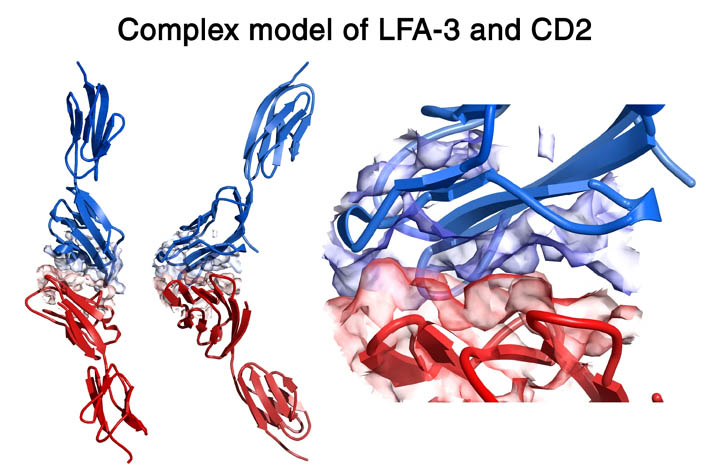
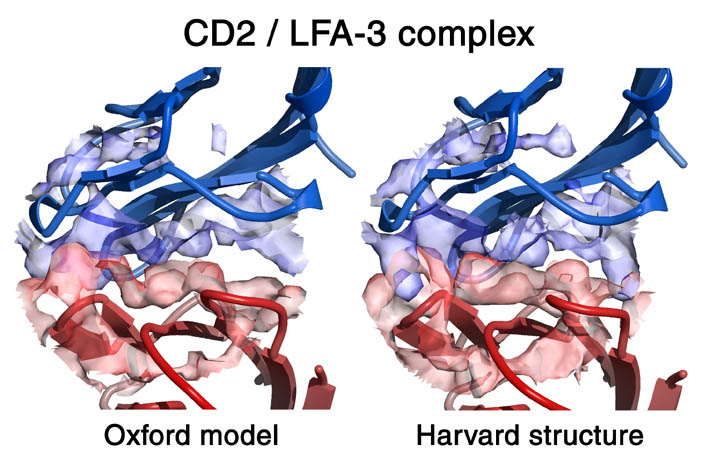
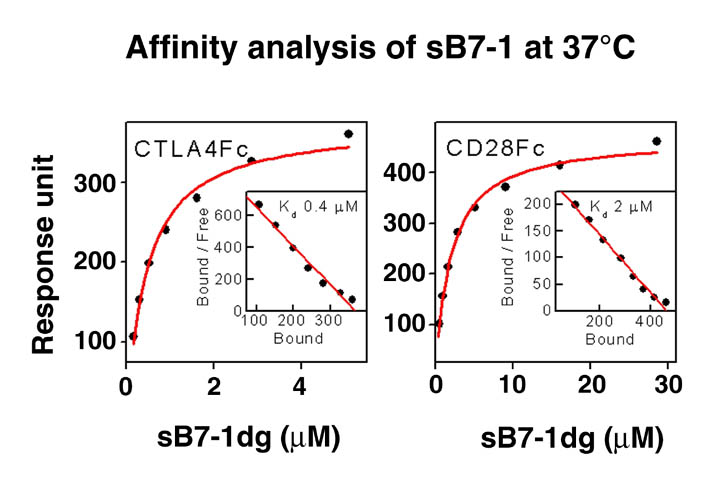
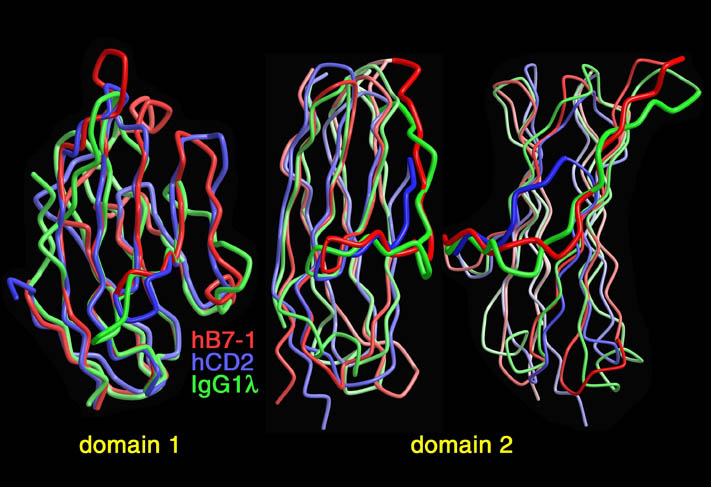
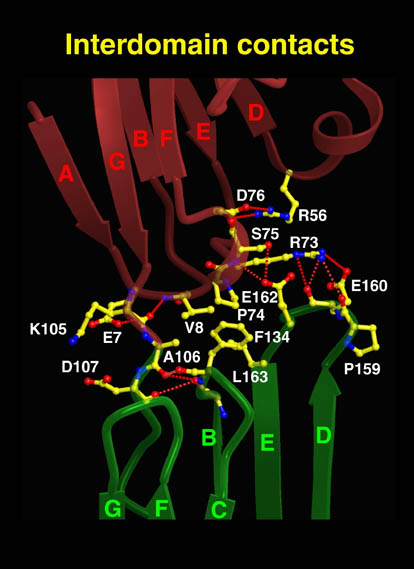
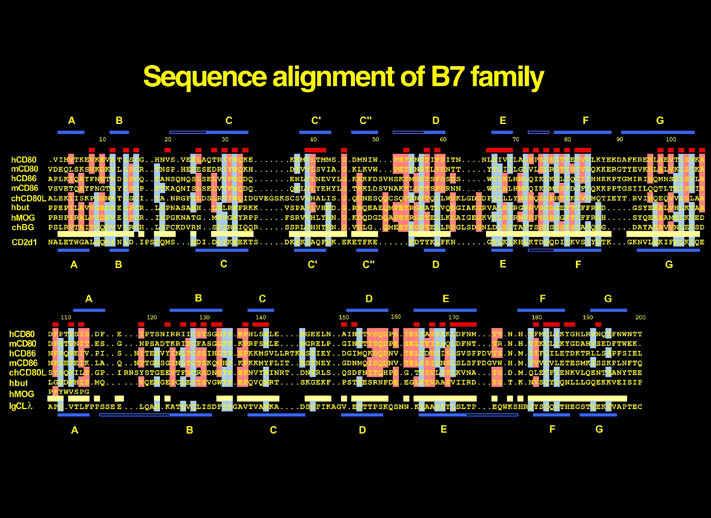
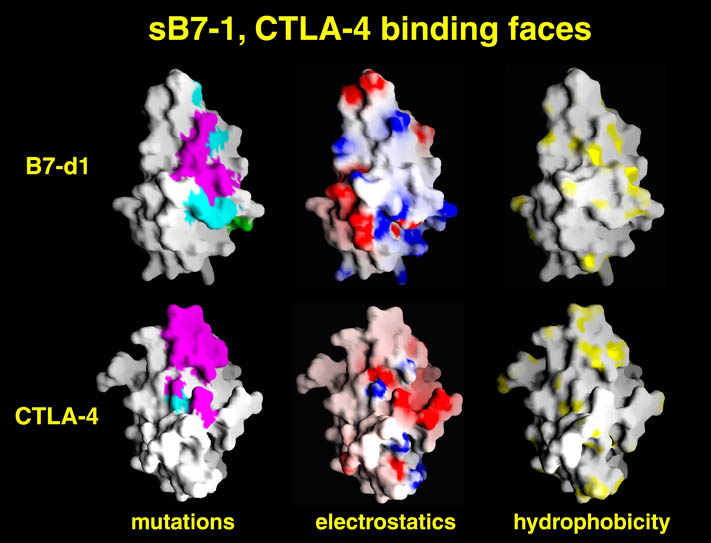
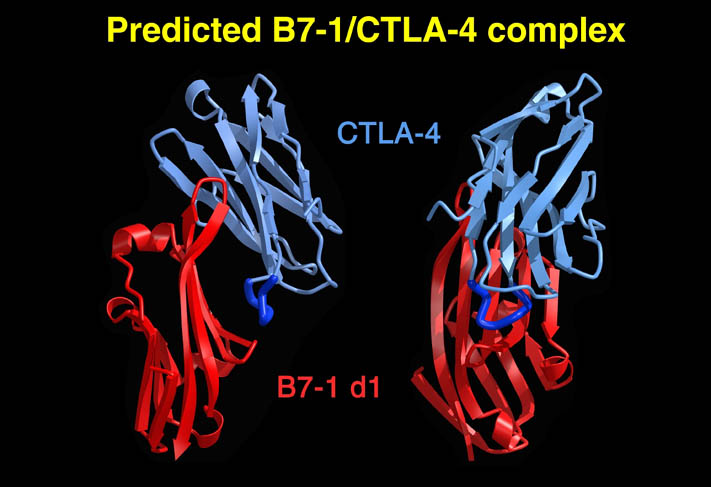
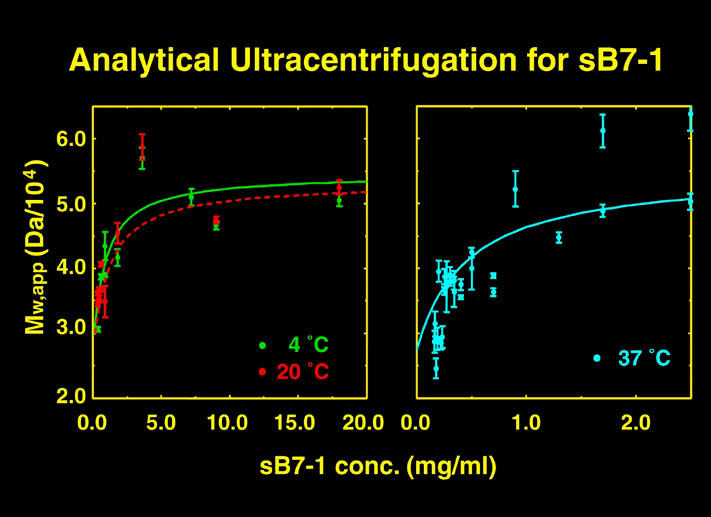
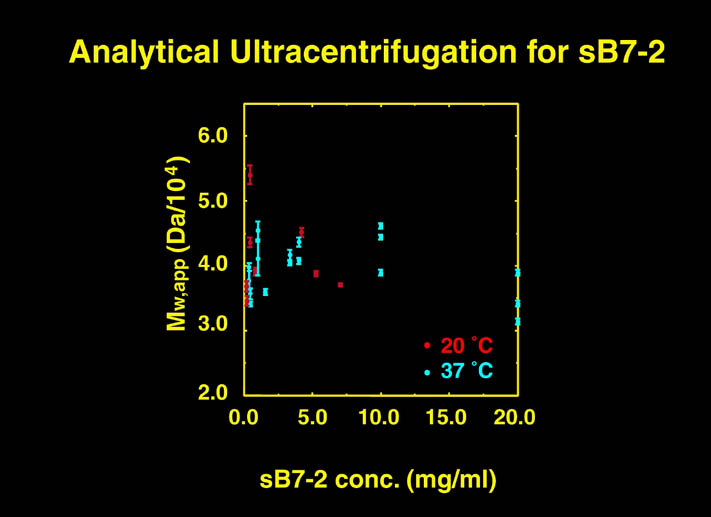
human LFA-3 d1・rat CD2 d2の可溶化型キメラ蛋白質の構造をrat CD2を用いた分子置換法により1.8A分解能で構造解析を行った。更に受容体であるCD2との複合体モデルを作成した。LFA-3/CD2複合体については、後にそれぞれの結合ドメインのみを用いた複合体の構造がHarvardのグループにより報告された。この構造は我々のモデルとほぼ同一であった。B7-1はSeMet置換体を用いて多波長異常分散法により3.0A分解能で構造解析を行い、CTLA-4を用いて複合体モデルを作成した。また結晶中でB7-1が二量体を形成していたことから、溶液中での二量体形成を超遠心分析(AUC)により確認した。B7-2についてもAUCによる実験を行ったが、二量体形成を見ることが出来なかった。
Stuart教授の将来計画としては、PRD1等のウィルス,ウィルス-受容体複合体,免疫系蛋白質の複合体,構造ゲノム等がある。彼は動物に感染するウィルスに最も興味がある。PRD1は脂質膜に覆われたウィルスで、この種のウィルスは未だ解析されていない。ウィルス-受容体複合体の研究では、ウィルスの細胞認識を明らかにすることによって、創薬へと結びつけることにある。免疫系蛋白質複合体の構造解析の目的は、蛋白質の分子認識についての知見を得ることと、それを基にした創薬にある。Polyhedrinというウィルス結合蛋白質がある。この蛋白質は微結晶しか得ることが出来ないが、キャピラリーにつめて測定すると2.0A分解能以上のパウダーパターンが得られる。重原子置換体についても同様2.0 A分解能で反射し、パウダーパターンに変化が見られる。これらのパウダーパターンのデータを用いた蛋白質の構造解析も行う。
オックスフォード大学構造生物学の将来
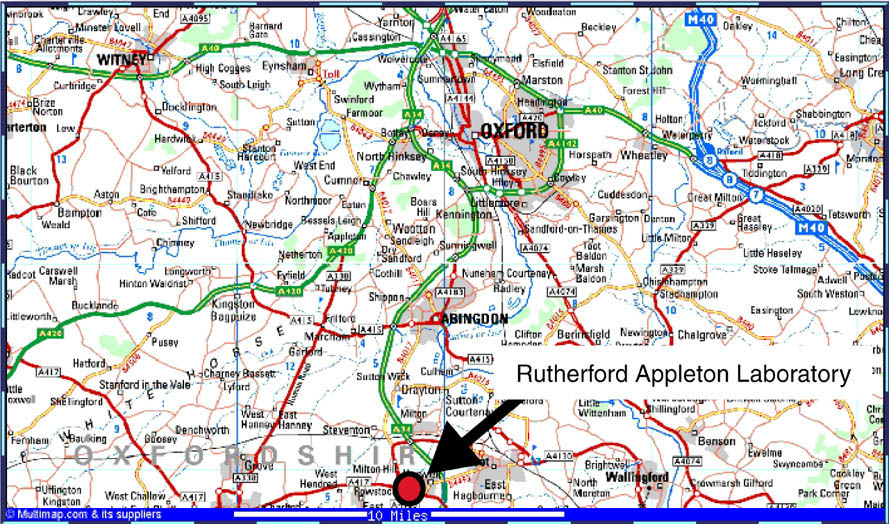
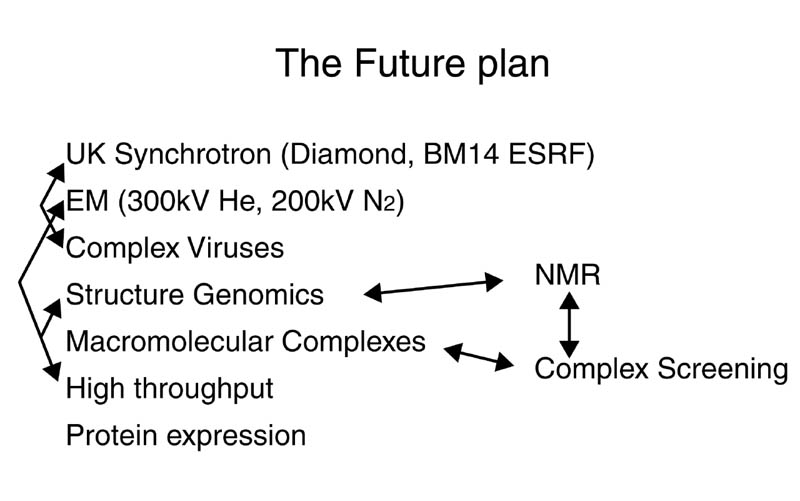
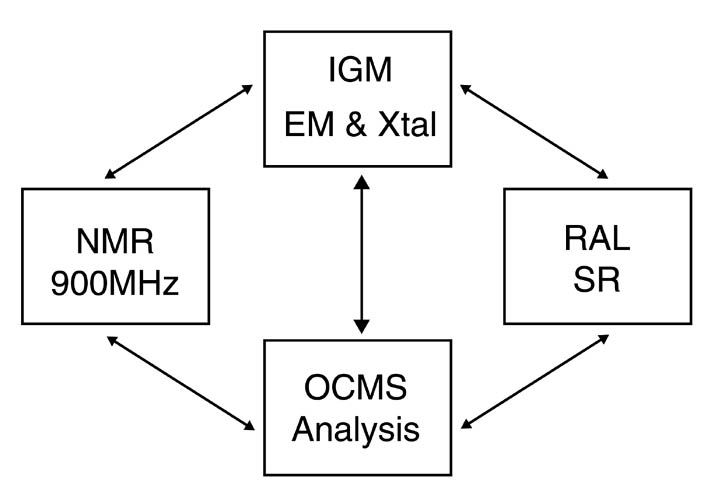
Daresburyに変わる次世代放射光施設DiamondがOxford近郊にあるRutherford Appleton Laboratoryに英国・フランス・財団Wellcome Trustの共同により建設されることが決定している。Diamondの設計はまだ行われていないが、5年後に稼動する予定である。DaresburyはDiamond稼動後2年間存続する予定で、7年後に運転を終了する予定である。Diamondは蛋白質結晶を主なターゲットにする予定で、蛋白質結晶ビームラインが6本出来ることは決定されている。当初計画されていたリングのエネルギーは3.0 GeVであったが、現在では3.5 GeVである。また英国では、ESRF BM14を買い取ることを計画している。ここではデータ測定だけでなくDiamondで導入される新技術(結晶の自動マウント等)の開発が行われると思われる。
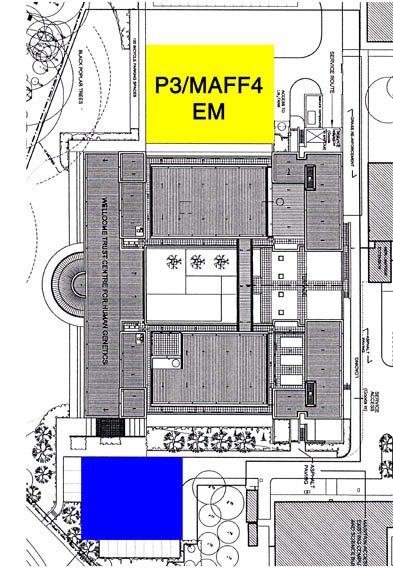
我々の研究所には、Facility for Evaluation of Infectious Particles (仮称)という施設が建設予定であり、この施設はP3(HIV, HBV等) /MAFF4 (FMDV, BTV等動物カテゴリー4)レベルのウィルスを扱うことが出来る。これにより全ての動物に対するウィルスを扱うことが可能になる。この施設には、300kV He, 200kV N2のEM2台,マススペクトル等が設置される。この様な施設の建設は英国内では初であり、外部にオープンし国際的な研究センターにする。
NMRでは、Ian Campbell 教授を中心にNational Centre for NMRの建設計画があり、900MHzの装置の導入を検討している。OCMSでは、マススペクトルやVIACore等の分析機器の充実が図られる。これらの全ての施設が密接に協力し、オックスフォード大学の構造生物学をより高いレベルに引上げる。
構造生物学の近未来
約10数年前に始まったヒューマンゲノムプロジェクトは、後1年程で全DNA配列の解析ができると言われている。ポストゲノムプロジェクトとして構造ゲノム(Structural Genomics)プロジェクトが近年盛んに議論されるようになった。構造ゲノムはバクテリア等のDNA配列を決定し、その中に含まれている蛋白質を順次構造解析を行い、生命現象を細胞又は生命体単位で捕らえるものである。日本・アメリカ等では既にプロジェクトが動き出しており、今後世界各国でプロジェクトが動き出すと思れる。英国に於いては、研究資金運営団体が構造ゲノムに集中的に予算を配分する計画を持っており、構造ゲノムプロジェクトが組織されることが予想される。現在の構造ゲノムもゲノムプロジェクトの初期同様にバクテリアを中心に行われている。しかし、最終的には構造ゲノムもゲノムプロジェクト同様、ヒューマンが主要なターゲットになると思われる。
ヒューマン構造ゲノムとバクテリア構造ゲノムの相違点は幾つか考えられる。
1)ヒューマンの方が、DNA中にコードされている蛋白質の数が多い(ヒューマンは 約10万種と言われているが、バクテリアは数千種)。
2)ヒューマンの蛋白質複合体の構造はより複雑である(原核生物のリボソームの沈降係数が70Sであるのに対して真核生物のものは80Sである)。
3)哺乳動物の蛋白質は、糖修飾を受けているものが多い。
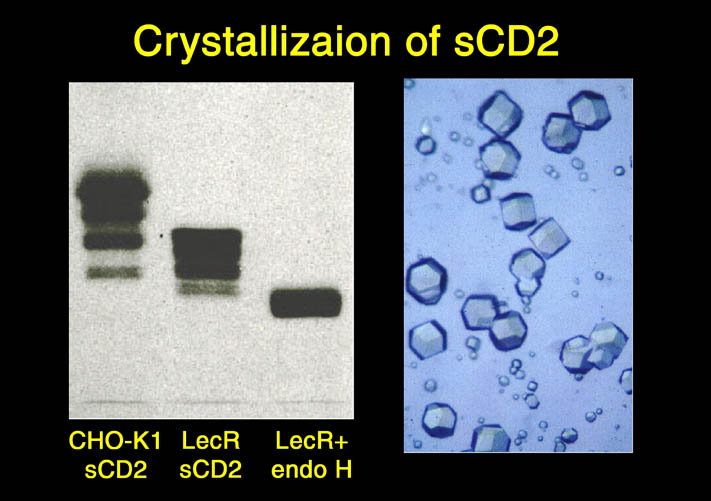
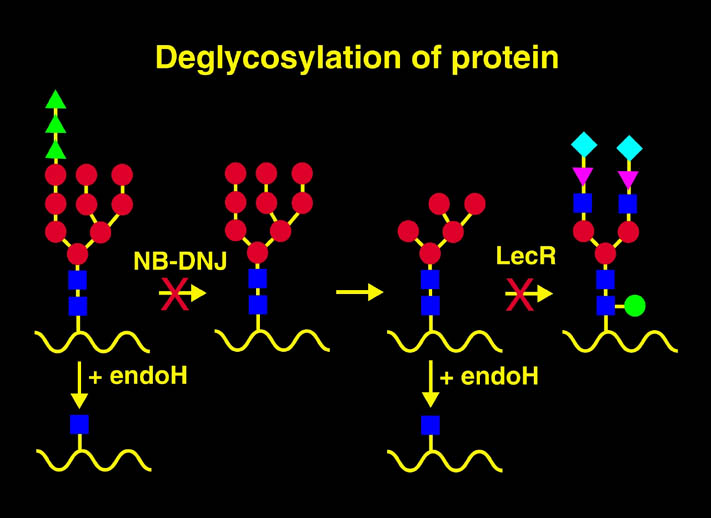
上記の様な相違点は、ヒューマン構造ゲノムプロジェクトの達成をより困難なものにする。3)については方法が確立されている。高マンノース型CHO細胞を使用することにより、蛋白質の糖修飾の複合型への修飾を抑えることができる。高マンノース型糖修飾蛋白質は、エンドHにより糖鎖一つをアスパラギン残基上に残して他の部分を分解することができる。
この様に得られた蛋白質は、結晶化に適している。蛋白質の取得で問題がありそうなものは、膜蛋白質やリボソーム等の複合体があげられる。これらの蛋白質発現系作成の技術開発は、今後行われていくと思われる。
スライドの図(1枚ずつ、じっくりと見たいとき、クリックしてください)
01 02 03 04 05 06 07 08 09 10
11 12 13 14 15 16 17 18 19 20
21 22 23 24 25 26 27 28 29 30
31 32 33 34 35 36 37 38 39 40
41 42 43 44 45 46 47 48 49 50
51 52 53 54 55 56 57 58 59 60
61
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}